 Photo 1. The larvae of the starfish, Asterina pectinifera. The length is
ca. 0.3 mm.
Photo 1. The larvae of the starfish, Asterina pectinifera. The length is
ca. 0.3 mm.
ワトソンとクリックがDNAの分子構造モデルを提案してから半世紀が経ちました. 20世紀後半の生物学は,このDNA分子構造の解明を起点として,分子遺伝学が大きく発展しました. ところが,セントラルドグマに表現される非常に単純化された遺伝情報の伝達モデルは,生物学に, 遺伝子に偏った研究と,遺伝子中心主義ともいえる状況も作り出してしまいました. R.ドーキンスの「利己的な遺伝子」はその象徴的産物であると言えます. また,ヒトゲノムプロジェクトが本格化すると, 生命保険会社は保険料の査定に保険加入者の遺伝情報を使うことを,いっせいに検討しはじめました.
進化論の世界においても,遺伝子中心主義は大きな影響を及ぼし,新ダーウィン主義が生まれました. すなわち,「まずDNAの塩基配列変化が無秩序に起こり, それがたんぱく質に翻訳されて個体に変化をもたらし,この変化のもたらす結果が, 生存競争に有利に働くと,そのDNAの変異が種内に蔓延し,進化をもたらす」と言うわけです. この考えは,上で述べた生命保険会社の行動の根拠ともなっています. また,「生存競争こそが生物進化の論理であるから, 人間社会も弱肉強食の生存競争が支配して当然である」 といった冷戦構造の崩壊以降強まった思想の背景も作り出しています.
ところが,当然のことながら遺伝子研究が進むと, このDNAが支配する生物の描像はあまりに単純化され過ぎていることが,次第に明らかになってきました. 進化論においては, 私たちは「生存競争は本当に進化の原動力なのか」という根本的な問いかけを行っています. 確かに,ダーウィンが見たガラパゴス諸島の生態系では, 少ない資源を奪い合い強者が生き残る厳しい生存競争が展開されていたのかもしれません. 私たちはこれを「貧困な世界」と呼んでいます. ところが,地球上で実際に圧倒的にたくさんの生物種が生活をしている熱帯雨林や海岸線に近い海は, 極めて豊かな環境にあり,生態系の食物網において一次生産者側の生物量が多く, 個体の消滅は飢餓ではなく,元気な個体も種間の捕食によって消滅するのが主と捉えられます. 逆に生存競争のもとでは,特定の生活条件のみが必須化し,生物種の多様性は生まれて来ないでしょう.
私たちは生物の主要な進化は,むしろ「豊かな世界」でこそ可能であったのではないかと考えています. 太田朋子氏は,無秩序に起こる様々の変異の多くは生物の生存に不利なものであることを明らかにしました (「分子進化のほぼ中立説」). けれども,厳しい生存競争のない豊かな世界では,個体はつねに飢餓に晒されているわけではなく, 体内にも蓄えを持ち,致死的な欠陥でない限り生存が許容されることになります. つまり,生物はぎりぎりの生活を強いられておらず,生活の仕方を変えることができます. 環境の変化に対応しても生活を適応させますが, 環境が変わらなくても内発的に多様な生活形態に可変なのです. 光合成の出現など,その最たる事例と捉えられます. だからこそ,地球に何度も起こった劇的な環境変化を乗り越えて, 生物はしぶとく進化し続けてきたのでしょう. このように,豊かな世界では,環境が許容する生活形が維持される限り, DNAの変異は蓄積されて行くことになります. まさにこのことが,1968年に木村資生氏が提唱した「分子進化の中立説」の内容であると言えます.
Photo 1. The larvae of the starfish, Asterina pectinifera. The length is
ca. 0.3 mm.
さて,私たちの研究対象であるヒトデは棘皮動物で,同じ後口動物ということでは, 私たちの遠い親戚に当たります. ヒトデの生息する海は,ヒトデにとって豊かな環境で,生存競争が支配する世界ではないことが, 私たちの実験から実際に明らかになりました. ヒトデはその発生において, ビピンナリア(Photo 1)とブラキオラリアという2段階の幼生期を経て成体に至ります. ところが,ヒトデの種類によっては,それらの双方とも,もしくは, いずれか一方の幼生期を経ずに成体に至るものがいます. これらのヒトデは卵が大きいという共通の特徴を持っています. 白井は,部分的に除去された卵の発生実験に基づき,ヒトデの住む豊かな環境では, 発生に必要な量を超えて栄養が卵に蓄積されていること(完全卵の14%でも発生可能)を明らかにしました. その結果,卵の余剰栄養の蓄積が幼生の退化を可能にしたと言う仮説を立てました. この考えは,より一般的には,余剰栄養蓄積による発生の異時性の進行に関する仮説でもあります.
脊椎動物では,もはや幼生形態はほとんど完全に退化してしまったと考えられます. “ほとんど”というのは,トリや爬虫類で卵の殻を裏打ちする構造や,哺乳類における胎盤など, 受精卵から派生するのにも関わらず成体には入り込まない細胞群は,見方によれば, 変態の名残と捉えられるからです. また,胚期と成体期ではヘモグロビンの型が異なるなど,分子種交代という現象があり, 生化学反応レベルでは,かつての幼生型が存在していると考えられます.
私たちは,ヒトデの卵に蓄積された栄養の量を実際に測るために熱測定を行いました. ヒトデ幼生は卵からの初期発生の段階では,捕食することができないので, 卵に蓄積された栄養によってのみ成長します. この栄養は,卵黄顆粒と呼ばれる粒子に蓄えられた複合たんぱく質です. これらのたんぱく質はエネルギー源であると同時に,体を作ってゆくための材料でもあります. したがって,エネルギー源としてのたんぱく質の量は実際にエネルギーに変換されて消費される量として計ら ねばなりません. これはまさしく幼生の発熱量で測られます.
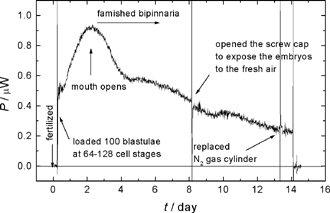 Fig. 1. Thermal power of the live larvae of Asterina pectinifera under
starved condition. A hundred eggs at the 64-128 cell stage were loaded 6 hours after fertilized at
the time 0.
Fig. 1. Thermal power of the live larvae of Asterina pectinifera under
starved condition. A hundred eggs at the 64-128 cell stage were loaded 6 hours after fertilized at
the time 0.
それでは,発生に必要な量を超えた過剰の栄養はどのようにすれば計測できるでしょうか. ヒトデの住む豊かな海では, ヒトデの幼生は消化器官が形成されると直ちに植物プランクトンを捕食するようになります. したがって,自然界ではヒトデの幼生は決して飢餓に陥ることはありません. しかし,私たちはあえて,幼生を絶食状態に置くことにしました. すなわち,ちょっと残酷かもしれませんが,卵から生まれた後,ずっと餌を与えないことにしました. すると,驚くべきことに,消化器官が形成された後, 3週間も餌なしで生き延びる幼生がいることが分かりました. 幼生の形態は単純化して行き,飢餓状況を示しますが,摂餌を開始すると, また生長を続け変態へも移行するのです.発熱量の測定は,この飢餓の間, 幼生はけっして活動を停止しているわけではないことを明らかにしています(Fig. 1). 実際に,卵から消化器官が形成されるまでの積算発熱量に比べて, 飢餓に陥ってからの積算発熱量のほうが何倍も大きいのです. つまり、発生は飢餓になるよりも十分にゆとりのある状況で進行しているのです.
私たちが今回測定したのは,直径約0.2 mmの小さい卵から2段階の幼生を経て発生するイトマキヒトデでした. この比較的小さな卵の間接発生型のヒトデにおいても,発生に必要な量をはるかに超えて, 栄養が卵に蓄積されていることを実証することができました.
Copyright © Research Center for Structural Thermodynamics, Graduate School of Science, Osaka University. All rights reserved.