 Photo 1. Prenauplius larva of Artemia in umbrella state.
Photo 1. Prenauplius larva of Artemia in umbrella state.
昨年の本レポートで,伝導型ミクロ熱量計(Thermometric社,LKB-2277/2277-201)の改良により,ヒトデの初期発生の発熱を長時間安定に観測できるようになったことを報告しました. その後1年間で装置の改良がさらに進み,より高感度の測定が可能になりました. その結果,直径わずか0.22 mmのアルテミア乾燥卵からの発生を,たった1個体の試料でも測定できるようになりましたので,紹介します.
生物の発生は,一般的に個体ごとに個性があって同調して進行しません. 実際,そのことが種の保存のために,重要な役割を果たしています. したがって,熱測定では1個体ごとの高感度観測が求められます. 一方,いったん始まった発生は途中で停止させておくことができないので,試料を熱量計に投入後,可能な限り速やかに測定が開始されることが望まれます. このことは,高感度測定になるほど難しくなります. これらの相反する要求を満たすため,熱量計に次の改良が施されました.
アルテミアは,エビやカニの仲間の甲殻類で,進化の系統樹で人間からは遠い存在です. 受精後卵巣内で発生が進行し,卵生の場合には,原腸胚期で発生がいったん停止した状態で放出されます. この卵を乾燥させると耐久卵(cyst)となり,無酸素状態で10年も保存可能になるという興味深い性質を持っています. この耐久卵には総重量の15%にも及ぶトレハロースが蓄積されていることが知られています. このため,卵は自らをガラス状態において休眠していると考えられます. また,アルテミアは内陸の塩湖に生息しています. 今回の実験に使用したアルテミアは,ユタ州のGreat Salt Lake産で,25%という飽和に近い高塩濃度で生息しています. そのため高度好塩性細菌とともに,アルテミアは非常に高い浸透圧の下で生活していることになります. 彼らはこの環境で生きてゆくために,細胞内に流れ込んでくるナトリウムイオンを効率よく排出するポンプを進化させてきたと考えられます.
Photo 1. Prenauplius larva of Artemia in umbrella state.
 Photo 2. Nauplius larva of Artemia at the second stage.
Photo 2. Nauplius larva of Artemia at the second stage.
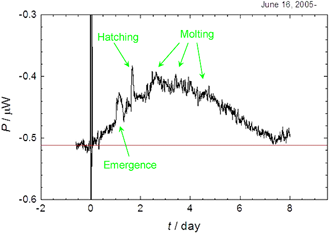 Fig. 1. Thermal dissipation of a nauplius larva of Artemia.
Fig. 1. Thermal dissipation of a nauplius larva of Artemia.
このように環境はたいへん厳しいものですが,逆に湖には,もはやアルテミアを捕食するような魚は居らず,またアルテミアの餌となる細菌や藻は豊富なので,アルテミアにとってみれば住めば都であるとも言えます. アルテミアの生活を制約する主な要因は,雨季に流れ込む淡水による表層の塩濃度の希釈,冬の水温の低下など,環境の季節変動です. 同じく季節変動と生活史の関係がよく研究されて来たのは昆虫ですが,昆虫の場合には捕食者との関係が常に考慮されねばならないので,捕食者のいないアルテミアは実に良い研究材料であると考えられます. なお,アルテミアの耐久卵は,金魚屋さんで熱帯魚の餌として安く手に入れることができます.
さて,本研究では,このように非常に興味深い生物であるアルテミアの耐久卵からの発生を,我々の改良された熱量計を使って観測しました. 乾燥耐久卵1個を0.1 mLの人工海水とともにガラス製アンプルに封じ込みます.アンプル内には3.2 mLの空気があり,アルテミアに十分の酸素が供給されます. 熱量計は20 °Cに保たれています. 海水に浸して約1日で,耐久卵の殻が割れて幼生が頭を出します. これをemergenceと言います. さらに,半日くらいかけて幼生は殻の外に出てきますが,まだ卵膜に覆われた状態にあります. この状態をumbrella stateと呼びます(Photo 1). やがて,卵膜を触角で突き破ってnauplius幼生が泳ぎ出てきます. これを孵化(hatching)と呼びます. その後,脱皮を繰り返しながらnauplius幼生は大きくなってゆきます(Photo 2). 捕食するようになるのは,第3脱皮が終わったあとになります. それまでの間は,母親からもらった卵黄のエネルギーを使って発生が進行します. 今回の実験では,アンプルに全く餌を入れていませんので,第3脱皮以降は飢餓状態になります.
発生の進行に伴う発熱量の変化の一例をFig. 1に示します. 1.14日と1.68日に鋭いピークが観測されます. これらはちょうどemergenceとhatchingに対応します. さらに,第1脱皮,第2脱皮,第3脱皮の各段階に緩やかな山が現れます. このように,熱測定によって,断続的に進行する発生の各段階の熱発生を追うことに成功しました. 約7.5日で発熱がなくなるのは,幼生が餓死したことによります. 昨年の本レポートで紹介したヒトデ幼生と違って,捕食可能になった幼生が僅か3日で餓死に至ることは,この動物の生活史を考える上で重要なヒントを与えています.
Y. Nagano, J. Thermal Anal. Calor. (投稿中).
Copyright © Research Center for Structural Thermodynamics, Graduate School of Science, Osaka University. All rights reserved.