 Photo 1. (Click to enlarge.)
Embryos of Aterina Pectinifera in the
24 cell stage.
Photo 1. (Click to enlarge.)
Embryos of Aterina Pectinifera in the
24 cell stage.
私たちは,現代生物学の主流となっているセントラルドグマに依拠した分子生物学的アプローチに対して,作業物質に依存しない事をその本質的特徴とする熱力学的物質理解の方法によって,分子の世界より上位の階層にある生命現象を捉えたいという願望をもっています. そのために私たちは,あえて生体関連物質の化学には手を出さず,複雑な生きた生物そのものを対象にして熱測定を試みてきました. 細胞分裂は,細胞が「生きている」ことを象徴的に示す最も神秘的な現象です. 顕微鏡を覗いて細胞分裂を観察するときには感動さえ覚えるものです. 今,生命科学は驚くべき速度で,細胞分裂のメカニズムをも解明しつつありますが,それでもなお実際の細胞分裂を目の当たりにした時,私たちの知りえたことはほんのわずかの断片に過ぎないことを実感させられてしまいます. むしろ分子論的メカニズムの解明が進むほど,逆に果たしてそういうアプローチで本当に細胞分裂の全体あるいは本質を捉えることができるのか,とさえ思えてきます.
Photo 1. (Click to enlarge.)
Embryos of Aterina Pectinifera in the
24 cell stage.
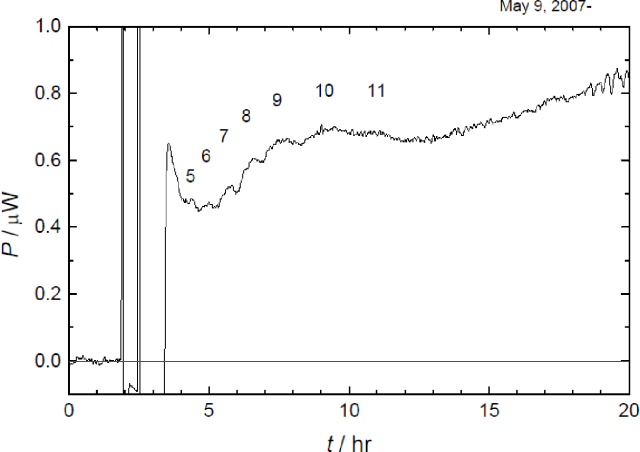
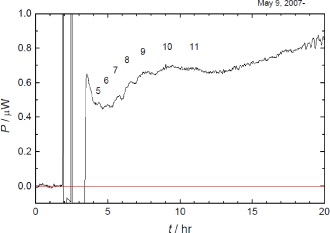 Fig. 1. (Click to enlarge.)
Thermogenesis of the eggs. Matured eggs were fertilized at t = 0, and 87 eggs were loaded in the microcalorimeter at
t = 3 hr. Sequential peaks were attributed
to the fifth to the eleventh cleavage.
Fig. 1. (Click to enlarge.)
Thermogenesis of the eggs. Matured eggs were fertilized at t = 0, and 87 eggs were loaded in the microcalorimeter at
t = 3 hr. Sequential peaks were attributed
to the fifth to the eleventh cleavage.
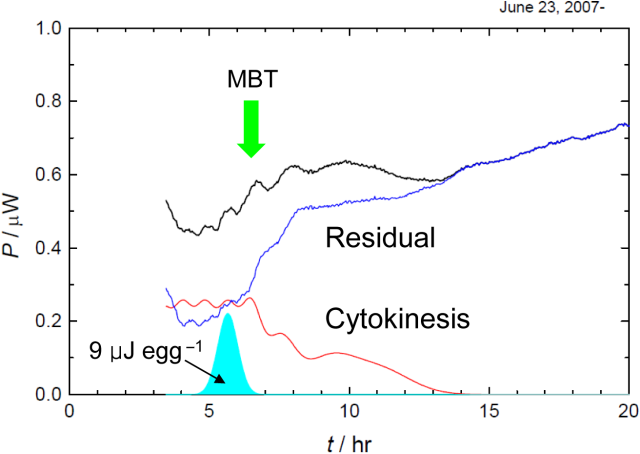
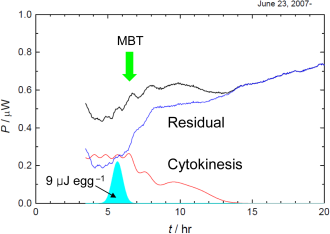 Fig. 2. (Click to enlarge.)
Heat of cytokinesis was tentatively separated. The residual shows an onset
in the vicinity of the 28 cell stage, in which
mid blastula transition (MBT) takes places.
Fig. 2. (Click to enlarge.)
Heat of cytokinesis was tentatively separated. The residual shows an onset
in the vicinity of the 28 cell stage, in which
mid blastula transition (MBT) takes places.
さて,受精卵の卵割は,細胞分裂を最もシンプルな形で私たちに見せてくれます. すなわち,初期発生における細胞周期は,交互に進行する染色体の複製(S期)と細胞質分裂(M期)のみから成っており,中期胞胚転移に到ってはじめて,DNAを読み取って,自らの遺伝情報に基づくたんぱく質を作り出すとされています. 高等学校の教科書にも取り上げられているウニと同じ棘皮動物のイトマキヒトデの初期発生では,受精卵は全割かつ等割の卵割を11回繰り返して,すなわち211細胞から成る中空球状の胞胚に成ります(Photo 1). 私たちは,いっせいに人工受精させたイトマキヒトデ卵の中から同調して卵割を始めた卵を約80個拾い集め,200 μLの人工海水に投入して,熱測定を行いました. すると,卵は卵割の周期に合わせた発熱の変動を示すことがわかりました(Fig. 1). さらに,並行した顕微鏡による卵割の観察から,観測されたそれぞれの発熱の山は第5卵割から第11卵割に相当することが分かりました. もし仮に,これらの発熱の山が染色体の複製によるものであるとすると,染色体の数は細胞の数と同じだけ増えていくので,2N倍の速度で発熱は大きくならねばなりません. しかし,細胞質分裂による発熱であるとすると,卵の細胞質の全量は不変であるので,発熱の大きさは一定であっても良いでしょう. 実際は後者に近いと考えられるので,観測された7つの発熱の山は細胞質分裂によるものであると結論されます.
このように,私たちは細胞質分裂に帰属できる発熱を初めて観測することに成功しました. 次に,全発熱から細胞質分裂の発熱を分離することを試みました. ただし,細胞質分裂の発熱が不変で,それぞれの発熱の山をガウス関数で近似できるとしました(Fig. 2). また,第9卵割以降は,細胞周期も長くなり,次第に細胞質分裂の同調性が失われてゆくことが知られており,実際に観測された山も緩やかになっています. このため,第9卵割以降はガウス関数の線幅を大きくすることにしました. その結果,1回の細胞質分裂で放出される熱は1卵あたりおよそ9 μJであることが分かりました. また,細胞質分裂による発熱を差し引いた残りの発熱は,第8卵割付近で急に立ち上がっており,細胞接着に関与するたんぱく質の発現もこのあたりで起ることから,この立ち上がりが中期胞胚転移に相当すると結論されました. ところで,細胞分裂には有糸分裂紡錘体と収縮環と呼ばれる細胞骨格の形成と運動が関与しています. 収縮環が細胞を引きちぎる力は,数10 nN程度であるとされています. しかし,卵の直径はおよそ0.2 mmに過ぎないので,収縮環がなす仕事は細胞質分裂の発熱と比べて無視しうるほど小さいことがわかります. 今回の熱測定の結果は,これまでの生物学によって明らかにされてきた分子機構が,細胞質分裂を本当にどの程度説明しているのかを評価するための最も基本的な材料を提供していると言えるでしょう.
長野八久,白井浩子,第43回熱測定討論会(札幌),3B1100 (2007).
Copyright © Research Center for Structural Thermodynamics, Graduate School of Science, Osaka University. All rights reserved.{kind=link}
{kind=link}
{kind=link}