



尋媶徯夘
GENERAL
惗懱暘巕壔妛尋媶幒偼丄
"扤傕弌棃側偄偙偲傪傗傞丅扤傕傗傜側偄偙偲傪傗傞"
傪儌僢僩乕偵丄惗棟妶惈暔幙乮栻暔傗撆慺側偳偺奜棃惈偺壔崌暔乯偍傛傃惗懱掅暘巕乮帀幙傗儂儖儌儞側偳偺惗暔偑杮棃帩偭偰偄傞壔崌暔乯偺俁師尦揑側宍偍傛傃惗懱撪偵偍偗傞摥偒傪夝柧偡傞偙偲傪尋媶栚揑偲偟偰偄傑偡丅
傢傟傢傟偑摼堄偲偡傞偺偼丄NMR側偳偺婡婍暘愅偲壔妛崌惉偱偡偑丄偙傟傜偺庤朄傪嬱巊偟偰惗柦尰徾傪尨巕偺惛搙偱偲傜偊傞偨傔偺尋媶傪峴偭偰偄傑偡丅
嬶懱揑偵偼丄埲壓偺係偮偺暘栰偺尋媶偑恑峴偟偰偄傑偡丅
枌嶌梡暘巕
侾丄傾儞僼僅僥儕僔儞俛
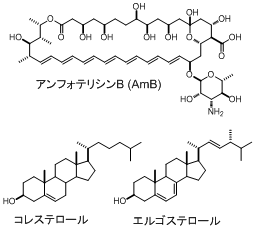 傾儞僼僅僥儕僔儞B乮AmB乯偼曻慄嬠Streptomyces nodosus偑惗嶻偡傞峈恀嬠嵻偱丄尰嵼偱傕廳撃側恀嬠姶愼徢偺帯椕偵峀偔梡偄傜傟偰偄傑偡丅
傾儞僼僅僥儕僔儞B乮AmB乯偼曻慄嬠Streptomyces nodosus偑惗嶻偡傞峈恀嬠嵻偱丄尰嵼偱傕廳撃側恀嬠姶愼徢偺帯椕偵峀偔梡偄傜傟偰偄傑偡丅
A倣B偼惗懱枌拞偺僗僥儘乕儖偍傕傃儕儞帀幙偲憡屳嶌梡偟丄僀僆儞僠儍僱儖暋崌懱傪宍惉偡傞偙偲偵傛偭偰妶惈傪敪尰偡傞偲偝傟偰偄傑偡丅
偦偺嵺丄AmB偼摦暔嵶朎枌偺僗僥儘乕儖偱偁傞僐儗僗僥儘乕儖傛傝傕丄僇價側偳偺恀嬠嵶朎枌偵偁傞僄儖僑僗僥儘乕儖偲嫮偔憡屳嶌梡偡傞偨傔丄恀嬠嵶朎枌拞偱僀僆儞僠儍僱儖傪宍惉偟傗偡偔丄慖戰撆惈偑敪婗偝傟傞偲峫偊傜傟偰偄傑偡丅
偟偐偟丄AmB偑僐儗僗僥儘乕儖偲僄儖僑僗僥儘乕儖偺旝柇側堘偄傪擣幆偡傞儊僇僯僘儉傗丄AmB偑宍惉偡傞僀僆儞僠儍僱儖暋崌懱偺峔憿偼偄傑偩偵偁偒傜偐偵側偭偰偄傑偣傫丅
偦偙偱丄AmB偑宍惉偡傞僀僆儞僠儍僱儖暋崌懱偺峔憿傪柧傜偐偵偡傞偙偲傪栚揑偲偟偰丄AmB偍傛傃僗僥儘乕儖傪19F傗13C偝傜偵2H側偳偱昗幆偟丄屌懱NMR應掕傪峴偄傑偟偨丅偙傟偵傛傝丄AmB傗僗僥儘乕儖偺帀幙枌拞偱偺塣摦惈傪柧傜偐偵偡傞偙偲偑偱偒丄AmB偲僄儖僑僗僥儘乕儖偑枌拞偱捈愙憡屳嶌梡偟偰偄傞偙偲偑柧傜偐偵側傝傑偟偨丅偝傜偵AmB摨巑傗AmB偲僄儖僑僗僥儘乕儖娫偺嫍棧應掕偵傕惉岟偟傑偟偨丅偙傟傜偺忣曬傪廤傔傞偙偲偱丄嵟廔揑偵暋崌懱偺峔憿寁嶼傪峴偄丄僠儍僱儖暋崌懱偺峔憿寛掕傪栚巜偟偰偄傑偡丅
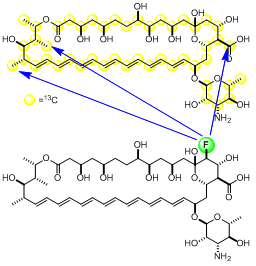
13C-AmB偲19F-AmB傪梡偄偨屌懱NMR偵傛傞暘巕娫嫍棧應掕
傑偨丄偙偺傛偆偵偟偰AmB僀僆儞僠儍僱儖暋崌懱偺峔憿偍傛傃嶌梡婡峔偑柧傜偐偵側傟偽丄傛傝暃嶌梡偺彮側偄栻嵻偺奐敪偵偮側偑傞偲婜懸偝傟傑偡丅
AmB僠儍僱儖偺悇掕峔憿
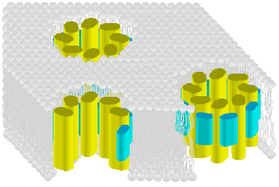
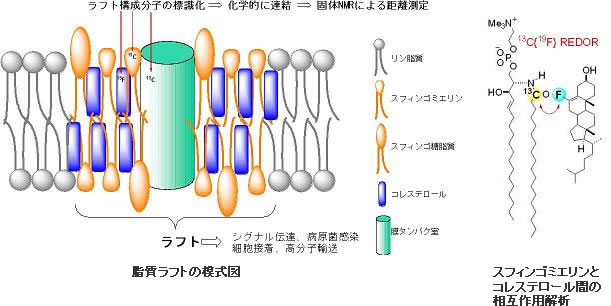
俀丄帀幙儔僼僩
惗懱枌偼寛偟偰嬒幙偱堦條側枌偱偼側偔丄晹暘晹暘偵傛偭偰惈幙傗峔惉惉暘偑堎側偭偰偄傞偙偲偑嵟嬤傢偐偭偰偒傑偟偨丅摿偵帀幙儔僼僩偲屇偽傟傞旝彫椞堟偼丄僗僼傿儞僑儈僄儕儞偲僐儗僗僥儘乕儖傪庡惉暘偲偟偰偍傝丄廃埻偺惗懱枌偲偼堎側傞惈幙傪帩偭偰偄傑偡丅偙偺帀幙儔僼僩偼丄僔僌僫儖揱払傗昦尨嬠偺姶愼側偳惗棟揑偵廳梫側摥偒傪偟偰偄傑偡偑丄暘巕娫偺憡屳嶌梡偵偮偄偰偼傎偲傫偳傢偐偭偰偄傑偣傫丅偦偙偱傢傟傢傟偼偙偺帀幙儔僼僩偵偍偗傞暘巕擣幆婡峔傪夝柧偡傞偨傔偵丄僗僼傿儞僑儈僄儕儞偍傛傃僐儗僗僥儘乕儖偺13C,19F,2H昗幆懱傪崌惉偟丄屌懱NMR偵傛偭偰暘巕娫嫍棧應掕偍傛傃塣摦惈偺夝愅傪峴偭偰偄傑偡丅偦偺寢壥丄僗僼傿儞僑儈僄儕儞偺帀幙枌拞偱偺岦偒傗塣摦惈丄偝傜偵暘巕娫憡屳嶌梡偵偮偄偰偺庬乆偺抦尒傪摼傞偙偲偑偱偒傑偟偨丅崱屻偼偙傟傜偺抦尒傪傕偲偵暘巕摦椡妛寁嶼傪峴偄丄儔僼僩宍惉偺儊僇僯僘儉傪柧傜偐偵偟偨偄偲巚偭偰偄傑偡丅傑偨丄尰嵼偼儔僼僩廤愊儁僾僠僪傗僞儞僷僋幙偲儔僼僩偲偺憡屳嶌梡夝愅偵岦偗偰丄僗僼傿儞僑儈僄儕儞偺寀岝昗幆懱傗岝恊榓昗幆懱偺崌惉傪峴偭偰偄傑偡丅傑偨丄巁惈嬨扽摐僔傾儖巁傪桳偡傞摐帀幙偱偁傞僈儞僌儕僆僔僪傕儔僼僩峔惉惉暘偲偟偰抦傜傟丄嵶朎娫偺愙拝傗憡屳嶌梡丄嵶嬠傗僂僀儖僗偺姶愼側偳偵娭楢偟偰偄傑偡丅偦偙偱丄僈儞僌儕僆僔僪偺帀幙儔僼僩撪偱偺嫇摦偵傕徟揰傪摉偰偰NMR傪梡偄偨尋媶傪恑傔偰偄傑偡丅
俁丄僶僀僙儖
僶僀僙儖偼傾儖僉儖嵔偺挿偄儕儞帀幙偲抁偄儕儞帀幙偐傜側傞僨傿僗僋忬偺帀幙夛崌懱偱偁傝丄挿嵔儕儞帀幙偑宍惉偡傞暯柺晹暘偼擇廳枌峔憿傪桳偟偰偄傞偙偲偐傜丄僶僀僙儖偼嵟彫偺帀幙擇廳枌儌僨儖偲峫偊傜傟偰偄傑偡丅抁嵔儕儞帀幙偵懳偡傞挿嵔儕儞帀幙偺妱崌偑懡偄僶僀僙儖偼奜晹帴応偵懳偟攝岦偡傞惈幙傪桳偟偰偄傑偡偑丄抁嵔儕儞帀幙偺妱崌偑戝偒偔側傞偲丄僨傿僗僋忬偺宍忬傪曐偭偨傑傑帴応偵懳偟偰摍曽揑偵懚嵼偟傑偡丅偦偙偱傢傟傢傟偼丄偙偺摍曽僶僀僙儖偵僒儕僲儅僀僔儞側偳偺枌嶌梡惈暘巕傪摫擖偟丄捠忢偺梟塼NMR應掕傪峴偄傑偟偨丅偦偺寢壥丄偙傟傜枌嶌梡惈暘巕偺枌拞偱偺峔憿偲懚嵼埵抲偺寛掕偵惉岟偟丄枌拞偱偺嶌梡敪尰婡峔傪柧傜偐偵偡傞偙偲偑偱偒傑偟偨丅傑偨嵟嬤偱偼僶僀僙儖傪梡偄偰忋弎偺帀幙儔僼僩傪嵞尰偱偒傞偙偲傪尒弌偟丄僗僼傿儞僑儈僄儕儞偺枌娐嫬壓偱偺峔憿夝愅側偳偵傕惉岟偟偰偄傑偡丅尰嵼偼ERATO偲嫟摨偟偰僶僀僙儖傪梡偄偨枌僞儞僷僋幙傗枌儁僾僠僪偺寢徎壔偵傕挧愴偟偰偄傑偡丅
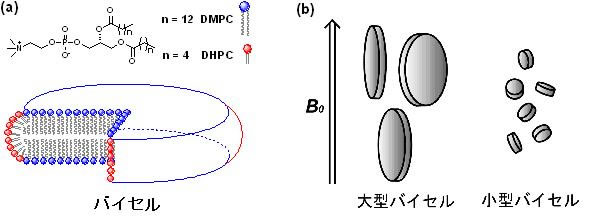
僶僀僙儖偺柾幃恾丅(a)挿嵔儕儞帀幙偼僶僀僙儖偺暯柺晹暘偱擇廳枌傪宍惉丅抁嵔儕儞帀幙偼僶僀僙儖偺僿儕偵懚嵼偡傞丅(b)戝宆僶僀僙儖偼帴応偵懳偟攝岦偡傞偑丄彫宆僶僀僙儖偼摍曽揑偵側傞丅
僶僀僙儖偵傛偭偰悇掕偟偨僀僆僲僼僅傾峈惗暔幙僒儕僲儅僀僔儞偺僇僠僆儞桝憲婡峔
係丄傾儞僼傿僕僲乕儖
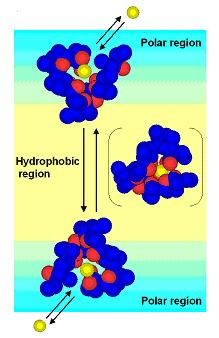
傾儞僼傿僕僲乕儖偼怉暔惈僾儔儞僋僩儞偱偁傞塓曏栄憯Amphidinium klebsii偑惗嶻偡傞儐僯乕僋側峔憿偺壔崌暔偱丄摿偵傾儞僼傿僕僲乕儖俁偼忋婰偺傾儞僼僅僥儕僔儞B傛傝傕嫮偄梟寣妶惈傗峈僇價妶惈傪帩偭偰偄傑偡丅偡偱偵傢傟傢傟偼傾儞僼傿僕僲乕儖俁偺愨懳棫懱攝抲傪娷傓慡棫懱峔憿傪寛掕偟偰偍傝丄偝傜偵妶惈敪尰婡峔偵偮偄偰傕尋媶傪峴偭偰偄傑偡丅偙傟傑偱偵丄偙偺壔崌暔偼惗懱枌偵嶌梡偟偰暋崌懱傪宍惉偟丄AmB傛傝傕戝偒側嵶岴乮僩儘僀僟儖宍偺嵶岴乯傪偁偗傞偙偲偱嫮偄妶惈傪帵偡偙偲傪柧傜偐偵偟傑偟偨丅傑偨儈僙儖傗忋弎偺僶僀僙儖傪梡偄偰枌拞偱偺傾儞僼傿僕僲乕儖俁偺棫懱攝嵗夝愅偵傕惉岟偟偰偄傑偡丅偝傜偵嬨廈戝妛戝愇嫵庼偺僌儖乕僾偲嫟摨偱丄傾儞僼傿僕僲乕儖俁偺晹暘峔憿傪崌惉偡傞偙偲偱棫懱壔妛偺掶惓傪偟傑偟偨丅尰嵼偼崅抦戝妛捗揷嫵庼偲嫟摨偱丄戝検攟梴偵傛傞傾儞僼傿僕僲乕儖俁偺嫙媼傪峴偄丄屌懱NMR傪梡偄偰傾儞僼傿僕僲乕儖偲僗僥儘乕儖偺憡屳嶌梡傪夝愅偡傞偙偲傪寁夋偟偰偄傑偡丅
揤慠暔崌惉
侾丄昗幆壔僶僼傿儘儅僀僔儞桿摫懱偺崌惉
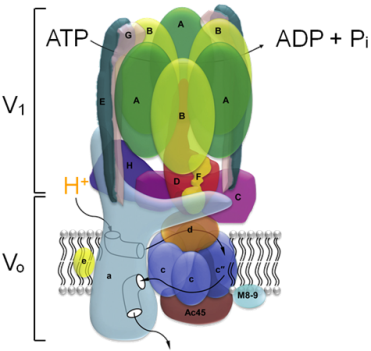
塼朎宆ATP傾乕僛乮V-ATPase乯偼恀妀惗暔偺撪枌偵峀偔懚嵼偡傞僾儘僩儞億儞僾偱偁傝丄僆儖僈僱儔偺撪晹傪巁惈偵曐偮栶妱傪扴偆堦曽偱丄崪慹闋徢側偳條乆側幘姵偵傕怺偔娭梌偟偰偄傑偡丅僶僼傿儘儅僀僔儞偼戙昞揑側V-ATPase慾奞嵻偱偁傝丄尰嵼丄V-ATPase偑娭梌偡傞條乆側惗棟婡擻傪夝柧偡傞偨傔偺暘巕僣乕儖偲偟偰斈梡偝傟偰偄傑偡丅偟偐偟側偑傜偦偺慾奞婡峔偵娭偡傞徻嵶側抦尒偼摼傜傟偰偍傜偢丄怴婯栻嵻奐敪偺娤揰偐傜傕偦偺暘巕儗儀儖偱偺嶌梡婡峔偺夝柧偑朷傑傟偰偄傑偡丅偦偙偱変乆偼丄僶僼傿儘儅僀僔儞偺19F, 13C昗幆懱傪崌惉偟丄屌懱NMR傪梡偄偨尨巕娫嫍棧應掕偵傛傝V-ATPase偲僶僼傿儘儅僀僔儞偺憡屳嶌梡傪夝愅偟傛偆偲峫偊偰偄傑偡丅
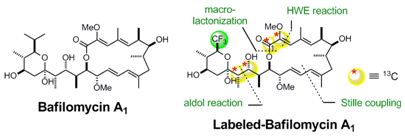
俀丄僗僺儘儕僪C偺慡崌惉
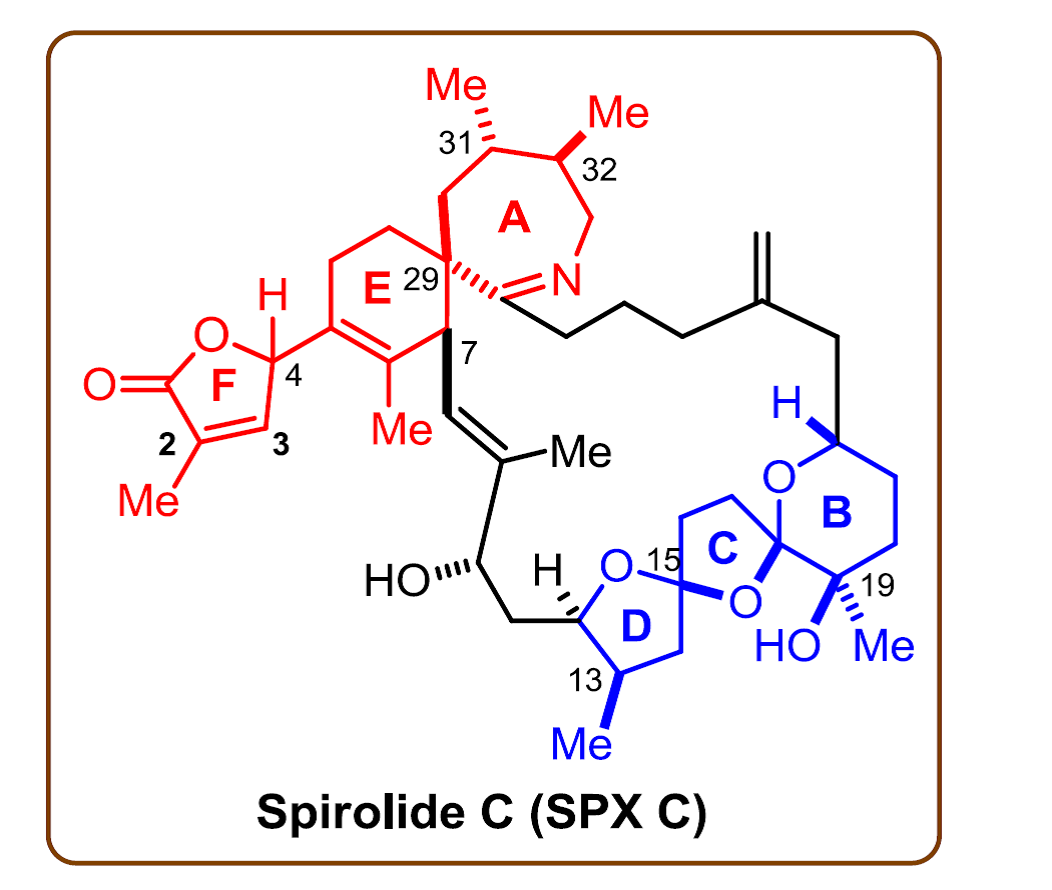 僗僺儘僀儈儞娐傪桳偡傞揤慠奀嶻撆偺1偮偱偁傞僗僺儘儕僪C (SPX C) 偼丄傾僙僠儖僐儕儞庴梕懱偵寢崌偟偰嫮椡側儅僂僗媫惈抳巰撆惈傪堷偒婲偙偡偙偲偑帵嵈偝傟偰偄傑偡丅偟偐偟側偑傜偦偺徻嵶側嶌梡婡峔偼偍傠偐枹偩偵慡愨懳棫懱攝抲丄摿偵4埵偵偮偄偰偼憡懳棫懱攝抲偝偊傕柧傜偐偵偝傟偰偄傑偣傫丅丂偙傟偼丄僗僺儘儕僪C偺峔憿揑摿挜丄偡側傢偪晄埨掕側γ-僽僥僲儕僪(F娐) 偑捈寢偟偨7,6-僗僺儘僀儈儞儐僯僢僩(A,E娐) 偲丄6,5,5-價僗僗僺儘働僞乕儖儐僯僢僩(B,C,D娐) 偑嵔忬扽慺偱楢寢偝傟偨暋嶨偱僼儗僉僔僽儖側峔憿傪桳偡傞偙偲偵婲場偟偰偄傞偲峫偊傜傟傑偡丅偦偙偱変乆偼丄僗僺儘儕僪C偺壔妛崌惉偵傛傞慡棫懱攝抲偺寛掕傪栚揑偲偟偰尋媶傪峴偭偰偄傑偡丅偝傜偵偼摨埵懱昗幆壔桿摫懱偺挷惢偵傛傞庴梕懱-僗僺儘儕僪暋崌懱峔憿偺暘巕儗儀儖偱偺夝愅傊偲揥奐偟傛偆偲峫偊偰偄傑偡丅
僗僺儘僀儈儞娐傪桳偡傞揤慠奀嶻撆偺1偮偱偁傞僗僺儘儕僪C (SPX C) 偼丄傾僙僠儖僐儕儞庴梕懱偵寢崌偟偰嫮椡側儅僂僗媫惈抳巰撆惈傪堷偒婲偙偡偙偲偑帵嵈偝傟偰偄傑偡丅偟偐偟側偑傜偦偺徻嵶側嶌梡婡峔偼偍傠偐枹偩偵慡愨懳棫懱攝抲丄摿偵4埵偵偮偄偰偼憡懳棫懱攝抲偝偊傕柧傜偐偵偝傟偰偄傑偣傫丅丂偙傟偼丄僗僺儘儕僪C偺峔憿揑摿挜丄偡側傢偪晄埨掕側γ-僽僥僲儕僪(F娐) 偑捈寢偟偨7,6-僗僺儘僀儈儞儐僯僢僩(A,E娐) 偲丄6,5,5-價僗僗僺儘働僞乕儖儐僯僢僩(B,C,D娐) 偑嵔忬扽慺偱楢寢偝傟偨暋嶨偱僼儗僉僔僽儖側峔憿傪桳偡傞偙偲偵婲場偟偰偄傞偲峫偊傜傟傑偡丅偦偙偱変乆偼丄僗僺儘儕僪C偺壔妛崌惉偵傛傞慡棫懱攝抲偺寛掕傪栚揑偲偟偰尋媶傪峴偭偰偄傑偡丅偝傜偵偼摨埵懱昗幆壔桿摫懱偺挷惢偵傛傞庴梕懱-僗僺儘儕僪暋崌懱峔憿偺暘巕儗儀儖偱偺夝愅傊偲揥奐偟傛偆偲峫偊偰偄傑偡丅
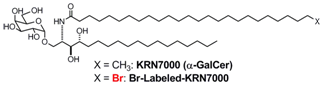
俁丄KRN7000 (α-僈儔僋僩僔儖僙儔儈僪) 偺崌惉
 KRN7000偼?-僈儔僋僩僔儖僙儔儈僪偲屇偽傟傞摐帀幙偱偁傝丄奀柸傛傝扨棧偝傟偨揤慠暔偺峔憿夵曄偵傛傝摼傜傟偨嫮椡側峈庮釃妶惈暔幙偱偡丅偦偺嶌梡婡峔偲偟偰丄柶塽嵶朎昞柺偵敪尰偡傞峈尨採帵僞儞僷僋幙CD1d偵寢崌偟丄僫僠儏儔儖僉儔乕T嵶朎昞柺偺TCR偲嶰幰暋崌懱傪宍惉偟偰庬乆偺僒僀僩僇僀儞嶻惗傪桿摫偡傞偙偲偑抦傜傟偰偄傑偡丅帀幙儕僈儞僪偱偁傞KRN7000偲CD1d偲偺嫟寢徎X慄峔憿夝愅偵傛傝偦偺寢崌條幃偑帵偝傟偰偄傞堦曽丄偙傟傪婎偵偟偨怴婯栻嵻偺崌惉尋媶偼丄偄傑偩桪傟偨栻嵻奐敪偵偄偨偭偰偄傑偣傫丅変乆偼丄偙偺棟桼偲偟偰嫟寢徎峔憿偺拞偱傕儕僈儞僪丄摿偵偦偺帀幙嵔偺揹巕枾搙偑掅偔惛枾側儕僈儞僪僨僓僀儞偑崲擄偱偁傞偨傔偲峫偊偰偄傑偡丅偦偙偱丄掅偄揹巕枾搙偱傕傛傝惓妋偵尨巕偺埵抲傪寛掕偱偒傞怴婯庤朄偺奐敪傪栚巜偟偰丄昗幆壔KRN7000偺崌惉傪偍偙側偭偰偄傑偡丅
KRN7000偼?-僈儔僋僩僔儖僙儔儈僪偲屇偽傟傞摐帀幙偱偁傝丄奀柸傛傝扨棧偝傟偨揤慠暔偺峔憿夵曄偵傛傝摼傜傟偨嫮椡側峈庮釃妶惈暔幙偱偡丅偦偺嶌梡婡峔偲偟偰丄柶塽嵶朎昞柺偵敪尰偡傞峈尨採帵僞儞僷僋幙CD1d偵寢崌偟丄僫僠儏儔儖僉儔乕T嵶朎昞柺偺TCR偲嶰幰暋崌懱傪宍惉偟偰庬乆偺僒僀僩僇僀儞嶻惗傪桿摫偡傞偙偲偑抦傜傟偰偄傑偡丅帀幙儕僈儞僪偱偁傞KRN7000偲CD1d偲偺嫟寢徎X慄峔憿夝愅偵傛傝偦偺寢崌條幃偑帵偝傟偰偄傞堦曽丄偙傟傪婎偵偟偨怴婯栻嵻偺崌惉尋媶偼丄偄傑偩桪傟偨栻嵻奐敪偵偄偨偭偰偄傑偣傫丅変乆偼丄偙偺棟桼偲偟偰嫟寢徎峔憿偺拞偱傕儕僈儞僪丄摿偵偦偺帀幙嵔偺揹巕枾搙偑掅偔惛枾側儕僈儞僪僨僓僀儞偑崲擄偱偁傞偨傔偲峫偊偰偄傑偡丅偦偙偱丄掅偄揹巕枾搙偱傕傛傝惓妋偵尨巕偺埵抲傪寛掕偱偒傞怴婯庤朄偺奐敪傪栚巜偟偰丄昗幆壔KRN7000偺崌惉傪偍偙側偭偰偄傑偡丅
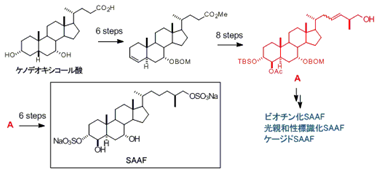
係丄儂儎惛巕妶惈壔桿堷暔幙乮SAAF乯偺崌惉
庴惛偺嵺丄惛巕偑棏偵憳嬾偡傞妋棪傪崅傔傞巇慻傒偺傂偲偮偲偟偰丄棏桼棃偺暔幙偑惛巕偺塣摦擻傪忋偘傞偲偄偭偨惛巕妶惈壔尰徾傗丄惛巕傪棏偺曽傊堷偒婑偣傞偲偄偭偨惛巕憱壔惈尰徾偑抦傜傟偰偄傑偡丅変乆偼僇僞儐僂儗僀儃儎偺棏巕偐傜偙偺椉妶惈傪桳偡傞暔幙偺扨棧偵惉岟偟乮嬐偐4 μg乯丄惛巕妶惈壔桿堷暔幙乮SAAF)偲柦柤偟傑偟偨丅傑偨丄NMR偍傛傃MS暘愅偵傛傝悇掕偟偨峔憿傪壔妛崌惉偵傛傝徹柧偡傞偲偲傕偵丄偦偺崌惉壔妛揑嫙媼傪幚尰偟傑偟偨丅尰嵼丄妋棫偟偨崌惉朄傪棙梡偟偰庬乆偺昗幆壔桿摫懱偺崌惉傪専摙偟偰偍傝丄SAAF偺妶惈敪尰婡峔偵偮偄偰傕尋媶傪恑傔偰偄傑偡丅