生物熱測定から何が分かるのか
もっぱら単純な化学物質の熱力学に携わってきたものにとって,生き物は巨大で複雑怪奇,平衡の熱力学は遠く及ばず,煮ても焼いても食えない代物でした(本当は煮ても焼いても「食えない」のは化学物質の方なのですが).
実際に,理論においても非線形非平衡の熱力学は未だほとんど闇の中,全く未開拓の分野と言ってよいでしょう.
それにも関わらず,少なからぬ研究者が果敢に,非平衡複雑系の最も魅力的な系として,生物に取り組んできました.
我々が専門とする熱測定においては,古くは A. L. Lavoisier
が18世紀に氷熱量計で,ねずみの発熱量を測定したのは良く知られています.
20世紀になり,E. Calvet
らが双子型ミクロ熱量計を開発すると,直ちに化学物質だけでなく動植物の熱測定が行われています.
応用面からも,発酵や土壌の研究に熱測定が利用されてきました.
しかし,熱測定は対象を問わず非常に安定に精度よく測れてしまうので,生物の熱測定例の蓄積もあるのですが,そのわりには,代謝研究における呼吸による酸素消費量との対応以外には,ほとんど注目されて来なかったのが実情です.
ところで,なぜ生物は発熱するのでしょうか.
その答えはすでに,E. Shrödinger によって与えられています.
すなわち,生物は非平衡で秩序状態を保ち続けているために,生きている限り,生成するエントロピーを排出し続けねばならないからです.
エントロピーを排出するには,高エントロピーの物質を排出するか,熱を出すかになります.
しかし,大きな自由エネルギーを取り出すためには,酸化などのエンタルピー支配の化学反応(発熱反応)を利用するのが有利であるので,発熱する生物が支配的になったと考えられます.
結果的に元気な生き物ほど熱いわけです.
もっとも,ごく初期の生命においては,吸熱する弱々しい生物もいたのかもしれません.
また,光を熱の一形態と見なせば,光合成は効率のよい熱機関とも言えます.
多くの生物では,糖や脂質の酸化で生成した ATP
をエネルギー輸送物質とし,そのエネルギーによって物質を移送したり化学反応を駆動しています.
おおざっぱに言って,生物の発熱は主にこれらの酸化反応(エネルギー代謝)に帰することができると考えられています.
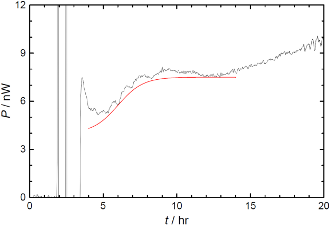 Fig. 1. (Click to enlarge.)
Early development of the starfish,
Asterina pectinifera at T
= 295 K. Mature eggs were fertilized at t = 0 hr.
87 synchronously cleaving eggs were loaded with 0.2 mL ASW in a 3.5 mL
ampoule. 7 peaks due to the 5th – 11th cleavages were observe from
t = 4 hr to 12 hr, while the peak at
t = 3.5 hr is instrumental. Thermal dissipation was
indicated as the value per embryo. An S-shape increase in the vicinity of
t = 6 hr was identified to
the mid-blastula transition.
Fig. 1. (Click to enlarge.)
Early development of the starfish,
Asterina pectinifera at T
= 295 K. Mature eggs were fertilized at t = 0 hr.
87 synchronously cleaving eggs were loaded with 0.2 mL ASW in a 3.5 mL
ampoule. 7 peaks due to the 5th – 11th cleavages were observe from
t = 4 hr to 12 hr, while the peak at
t = 3.5 hr is instrumental. Thermal dissipation was
indicated as the value per embryo. An S-shape increase in the vicinity of
t = 6 hr was identified to
the mid-blastula transition.
生物の発生を観察していて興味深いのは,生物は卵から大人になるまで,けっして緩やかに連続的に変化してゆくのではなく,形態変化や蛋白質の発現などから,発生の諸段階を階段的に転移してゆくように思われることです.
エネルギー・物質代謝(流れ)に支えられた安定な散逸構造が,突然に別の安定な構造に移ってゆくわけで,生物がカオスの縁にあることを特徴づけている現象であると理解されます.
昨年,長野らはイトマキヒトデの初期卵割において,細胞質分裂にともなう発熱を初めて観測したことを報告しました.
胚の全発熱から細胞質分裂の周期的発熱を取り除いてやると,第7から第8卵割にかけて,発熱が階段状に増えていることが分かります(Fig. 1).
ちょうどここが中期胞胚転移と呼ばれるところで,ウニの胚においても蛋白質合成に同様の立ち上がりが観測されています.
単に増加すると言うだけでなく,転移の後,原腸胚形成で再び増加するまで,発熱量が一定になっていることが大事な点です.
イトマキヒトデの卵成熟,受精においても同様の発熱のジャンプを観測し,エネルギー代謝の観点からも,発生が階段状に進行してゆくことが支持されます.
このような段階的な発生は,個体発生においてはすでに遺伝子上にプログラムされているので,決まった手はずで進んでゆきます.
しかし,これも進化の過程で獲得されてきたわけなので,何がこの転移を引き起こし,何によって次の発生段階が決められたのかを知る手がかりが,それぞれの発生段階のアトラクターを不安定化する要因として,どこかに潜んでいると思われます.
(長野 八久,白井 浩子)
Copyright © Research Center for Structural Thermodynamics,
Graduate School of Science, Osaka University. All rights reserved.
{kind=link}