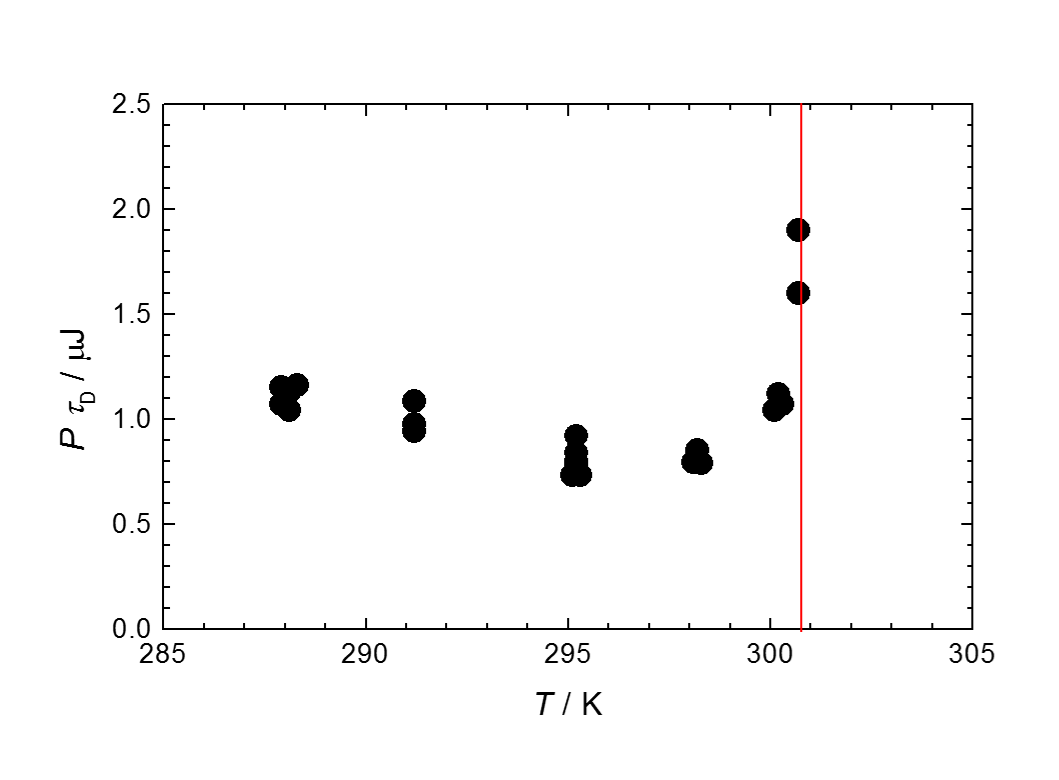
 Fig. 1. Energy consumption per cell of Dictyostelium discoideum Ax-2 during a doubling time in the growing phase. The line at T = 27.5 °C indicates the upper temperature limit of this organism.
Fig. 1. Energy consumption per cell of Dictyostelium discoideum Ax-2 during a doubling time in the growing phase. The line at T = 27.5 °C indicates the upper temperature limit of this organism.
Fig. 1. Energy consumption per cell of Dictyostelium discoideum Ax-2 during a doubling time in the growing phase. The line at T = 27.5 °C indicates the upper temperature limit of this organism.
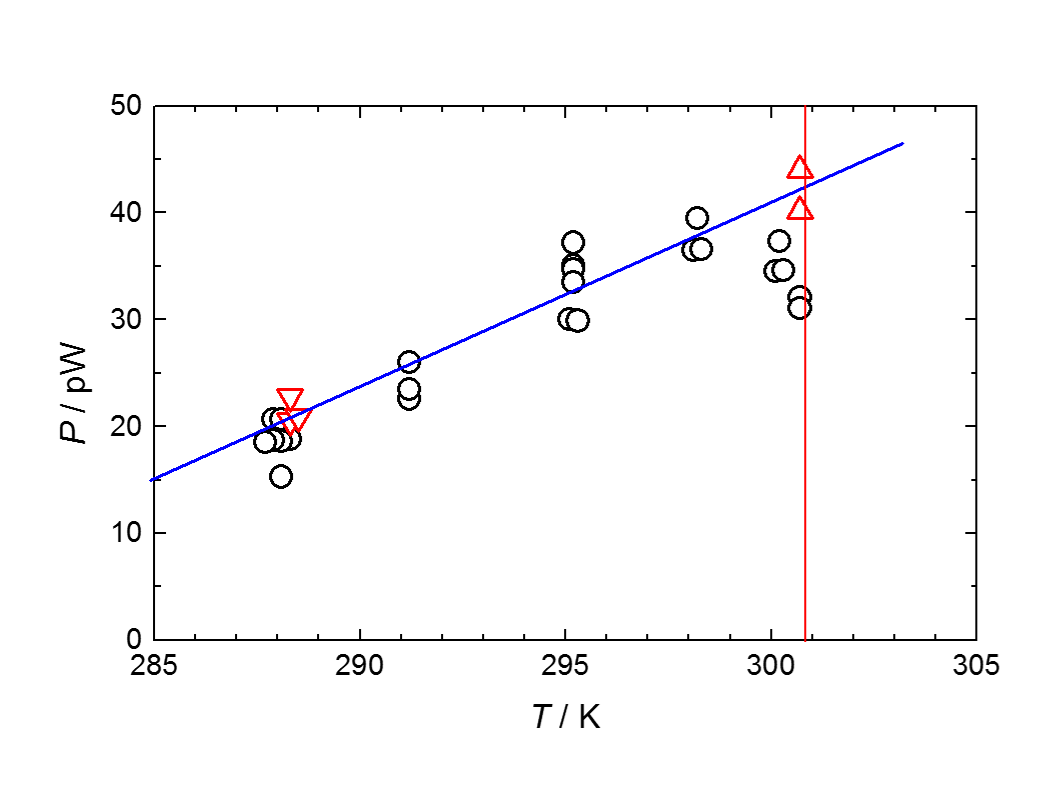 Fig. 2. Thermal dissipation rate per cell of Dictyostelium discoideum Ax-2. Triangles show those soon after temperature jumps from the optimum temperature.
Fig. 2. Thermal dissipation rate per cell of Dictyostelium discoideum Ax-2. Triangles show those soon after temperature jumps from the optimum temperature.
熱力学法則は、宇宙のすべての物質の運動を支配していることを主張しています。しかし、私たちは未だ宇宙の成り立ちすべてについて知っている訳ではないので、これは今なお大胆な主張です。他方、統計力学は、どのような性質をもつ作業物質なら、熱力学の諸法則を満たす運動をするのかを明らかにしています。実際には、私たちの身の回りは、すべて化学物質で構成されているので、世界を化学物質の集合体と見なす限りにおいて、人間も含めて熱力学法則の完全な支配の下にあります。
しかし、階層構造を成す宇宙の上部構造である生態系や社会における運動、相互作用に着目するならば、必ずしも熱力学法則が支配しているわけでないように見えます。例えば、人間関係において、熱力学第0法則を満たす熱力学温度を持ち込むことはできそうにはありません。これは、その階層における運動の担い手(agent)が、熱力学法則における作業物質の要件を満たしていないからです。Prigogineらによって発展させられた非平衡系の熱力学も、基本的には化学熱力学であるので、散逸構造の生成、安定性はエントロピーによって評価されています。したがって、非平衡熱力学における発展法則(基準)が厳密に適用されるのは、そもそも化学物質の階層に限られていることに注意する必要があります。 さて、生物も階層構造を成しています。その土台を成す代謝系は、化学反応系であるので、化学熱力学の完全な支配下にあります。それでは、生物個体全体はどうなのでしょうか。面白いことにZotinら(1967)は、動物の初期発生において、エネルギー代謝速度の変化が、個体レベルであっても、"エントロピー生成速度最小の原理"(より一般的には"普遍的発展基準")で表されることを示しました。このことは、生物個体の上部階層構造も、代謝系の化学熱力学に強く支配されていることを主張しています。しかし、生物個体は、単なる代謝系の流れの上に生成した散逸構造ではなく、それ自身が代謝系を制御する能動性を持っています。
生物が最適条件で生存しているときには、個体は代謝系を上手く手なずけて、代謝系の能力の範囲内において自由に生活しています。ところが、最適条件から外れてくると、生物個体の活動と代謝系の間に葛藤が発生することになります。適応限界を越えてしまうと、代謝系は個体の要求に応えられなくなり、最後は個体の死に至ります。したがって、この葛藤の役割は重要で、代謝系との葛藤があるからこそ、生物個体は、自分の生存を保障する条件の範囲内で活動するようになります。動物の初期発生で観察される最適温度において消費エネルギーが最小になるという現象も、熱力学法則の直接の現われと言うよりも、この葛藤の結果として現われていると考えられます。 より具体的に見ると、真核細胞において主なエネルギー代謝を担っているのは、ミトコンドリアという細胞小器官(organelle)です。酸素呼吸によって糖を分解し、ATPを生産します。ATPは細胞内に溜めておくことができないので、ATPを生産するミトコンドリアは、ちょうど需要に合わせて電力を供給する発電所のような役割を担っています。また、ミトコンドリアは細胞内共生(endosymbiosis)から進化した器官で、自分自身のDNAを保持し、2重膜構造を持っているのはその名残でもあります。ミトコンドリアの活動は、核の遺伝子の完全な制御の下にあるわけではなく、進化的に一定の独立性を保った共生の性格を強く帯びているのが面白いところです。エネルギー供給に関わる細胞内葛藤は、ミトコンドリアと宿主細胞の間で起こります。
細胞性粘菌Dictyostelium discoideumは、真核細胞生物であり、"社会的アメーバ"としてよく知られていますが、最適増殖条件下では単細胞アメーバとして非社会的に生活します。しかし、高温条件には弱く、適応限界温度27.5 °C以上では社会性を発揮することなく死滅します。最適温度は22 °Cとされているので、高温側の適応温度範囲は非常に狭いことになります。
増殖中の細胞の熱発生の主要な部分は、ATP生産とその消費によっているので、熱量測定は、完全非侵襲(non-invasive)のミトコンドリアの活動量評価法として有効です。Fig. 1は増殖期にある細胞性粘菌D. discoideum Ax-2、1細胞の1細胞周期あたりの総発熱量の温度変化を示しています。実際に細胞性粘菌においても、最適温度付近においてエネルギー消費が最小であることがわかります。しかし、実はこれは細胞周期が大きく温度変化し、最適温度で最小になることによるもので、単位時間当たりの発熱量は逆に25 °C付近で最大になっています(Fig. 2)。
一方、最適温度から温度ジャンプさせて、直後の発熱量を測ってやると、測定温度範囲内においては、発熱量はほぼ温度に比例していることが分かります。したがって、高温適応限界温度近くで培養された細胞で発熱量が減少するのは、ミトコンドリアの体積減少あるいは劣化が起こっているためと考えられます。すなわち、最適温度以上では、正常なミトコンドリアの複製が強く抑制されてしまっていることになります。細胞性粘菌の適応限界温度27.5 °Cは、代謝系自身の限界ではなく、個体との葛藤の結果として代謝系の縮小が起こっていることが示唆されます。
Copyright © Research Center for Structural Thermodynamics, Graduate School of Science, Osaka University. All rights reserved.