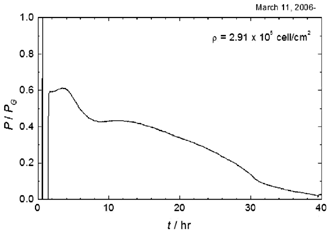
 Fig. 1. Thermogenesis of starved Dictyostelium discoideum Ax-2 at T = 295 K. Density of cell was 2.9×105 cell/cm2
at the starvation treatment, t = 0.
Fig. 1. Thermogenesis of starved Dictyostelium discoideum Ax-2 at T = 295 K. Density of cell was 2.9×105 cell/cm2
at the starvation treatment, t = 0.
長野らはこれまでに,ヒトデ幼生,アルテミア幼生の飢餓状態における熱測定を行い,卵黄の余剰蓄積とこれらの動物の生活史の関係について検討してきました. 実際に自然界においては,これらの動物は種の保存のために飢餓への対応が迫られるというようなことは無かったと考えられます. アルテミアの耐久卵(シスト)形成も,飢餓にではなく,低温,乾燥という環境変動への有効な対応となっています. 他方,地球上には飢餓への対応がプログラム化されている生物も存在します. その代表例が細胞性粘菌です. 栄養が豊富にある環境では,細胞性粘菌は単細胞のアメーバとして増殖し続けますが,飢餓に置かれると,飢餓を察知した細胞の中で,たまたま細胞周期の増殖・分化移行(GDT, Growth Differentiation Transition)点に先に到達した細胞からcAMP分泌などの分化形質を発現させ,細胞周期から離脱します. これらの細胞はcAMPに誘引されて集合体をつくります. 集合体の中では,予定胞子細胞と予定柄細胞への分化が起り,子実体を形成して,前者の細胞のみが胞子となって生き延びます. 興味深いことに,早くにGDT点に到達した細胞群が胞子となって生き延びるように運命づけられています.
細胞性粘菌ではGDT点で発現する形質が分化への移行に重要な役割を担っていますが,それはすべての細胞がGDT点を通過し細胞周期を離脱するまでに1細胞周期(8時間から10時間)掛かることも意味します.したがって,仮に細胞がGDT点を通過するまで増殖期の活動を多く維持しているならば,先にGDT点を通過した細胞ほど,細胞内に栄養を保存していることになり,それらの細胞が最終的に胞子となることを合理化するとも期待されます. しかし,実際には先にGDT点を通過して胞子になる細胞も,集合体形成後に細胞分裂を起こすので,結局飢餓に入って以降にすべての細胞が等しく1回分裂することも知られています. 確かに細胞性粘菌では飢餓によって分化が誘導されますが,はたして細胞の運命まで物質的・エネルギー的制約が関与しているのでしょうか. さらに,すべての多細胞生物に見られる生き延びる細胞(生殖細胞)と死すべき細胞(体細胞)への分化は,このような資源の枯渇によって引き起こされた進化なのでしょうか. 長野らはこの根本問題に迫るべく,細胞性粘菌の増殖・分化移行に伴う代謝量の変化を発熱量の変化として精度よく測ることに成功しました.
温度295 Kで静置培養された細胞性粘菌Dictyostelium discoideum Ax-2株は,一昨年の本レポートで紹介した熱量計試料容器(3.3 mLガラスアンプルにアルミ合金製のねじ蓋をつけたもの)に投入されます. 熱測定は,伝導型ミクロ熱量計(Thermometric社,LKB-2277/2277-201)を295 Kに設定して行われました. 細胞は熱量計の中で約1日培養され,発熱量から増殖期にある細胞の平均発熱量と倍化時間が決められます. この後,アンプル内の栄養培地をリン酸緩衝液(pH=6.5)に置換して飢餓処理を行います. 続いてリン酸緩衝液を吸引し,再び熱量計に投入して熱測定を継続します. 細胞は飢餓処理後アンプル内で集合し,子実体の形成にまで至ります. 子実体形成は,熱測定終了後,試料容器を洗浄したときに,洗液に胞子が観察されることで確かめられます.
Fig. 1. Thermogenesis of starved Dictyostelium discoideum Ax-2 at T = 295 K. Density of cell was 2.9×105 cell/cm2
at the starvation treatment, t = 0.
先ず増殖期における平均発熱量は,上述の培養条件において,22 pW/cellであり,倍化時間は9.5時間であることが明らかになりました. また,発熱量は飢餓処理後直ちに増殖期の60%以下になりました. さらに,飢餓処理後,細胞は時間と共に単調に発熱量を減衰させてゆくのではないことも明らかとなりました. Fig. 1は飢餓処理後の発熱量Pの変化を飢餓処理直前の発熱量PGで規格化したものを示します. 発熱は飢餓処理後も緩やかですが増加し続け,4時間でピークに達します. その後急な減衰に転じて飢餓処理後8時間でP/PGは0.42程度になります. これは,飢餓処理後も細胞周期と増殖期の活動が,GDT点に達するまでの間,部分的に維持されているとして説明されます. この後再び発熱は緩やかに上昇し,飢餓処理後約12時間に2つ目のなだらかなピークを形成し,これ以後発熱は減少してゆきます. 2つ目のピークは,細胞が集合してマウンドの形成を終わり予定柄細胞と予定胞子細胞への分化を始める時期と一致しています. この様に,発熱量の時間変化と増殖・分化転移の一連の過程がよく対応していることが明らかになりました.
長野八久,桑山秀一,日本発生生物学会第39回大会(広島),2B1030 (2006).
Copyright © Research Center for Structural Thermodynamics, Graduate School of Science, Osaka University. All rights reserved.